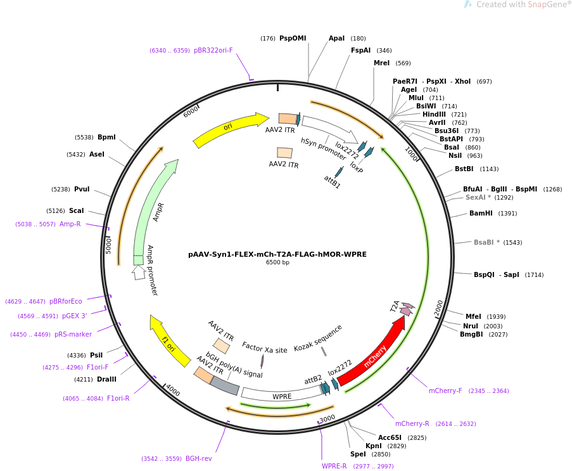
Random, hopefully Useful stuff that we have developed in the lab. Some published, some not...
|
opioids
enkephalin dynorphin DAMGO oxymorphone naloxone |
other neuropeptides
substance P gastrin-releasing peptide oxytocin cholecystokinin |
Name |
Caged variant of: |
Wavelength sensitivity |
Mu EC50 (LE=90 nM, D8=63 nM, DAMGO=1.5nM) |
Delta EC50 (LE=3 nM, D8=10 nM) |
Kappa EC50 (LE inactive, D8=7 nM) |
Photolysis kinetics (approx) |
Available through NDSP? |
Reference |
CYLE |
leu-enkephalin |
355 nm |
16 µM |
1.7 µM |
inactive |
10's of µs |
yes |
|
CYD8 |
dynorphin 8 |
355 nm |
23 µM |
3.9 µM |
16 µM |
10's of µs |
yes |
|
CNV-Y-LE |
leu-enkephalin |
355-405 nm |
12 µM |
0.4 µM |
inactive |
10's of µs |
yes |
|
N-MNVOC-LE |
leu-enkephalin |
355-405 nm |
34 µM |
17 µM |
inactive |
hundreds of ms |
no |
|
N-MNVOC-D8 |
dynorphin 8 |
355-405 nm |
41 µM |
28 µM |
33 µM |
hundreds of ms |
no |
unpublished |
CNV-Y-DAMGO |
DAMGO |
355-405 nm |
1.7 µM |
N.D. |
N.D. |
10's of µs |
no |
|
PhOX |
oxymorphone |
355-405 nm |
>100 µM |
>100 µM |
>100 µM |
10's of µs |
no |
|
PhNX |
naloxone |
355-405 nm |
>100 µM |
>100 µM |
>100 µM |
10's of µs |
no |